2024年2月11日の「前胸腺抑制神経ペプチド(1)」で、前胸腺抑制神経ペプチド、ミオサプレッシンの発見について紹介した。最後に『「ミオサプレッシン構造決定後」と「ミオサプレッシン研究から派生した成果」について続編を書くことにする。』と書いておきながら約10ヶ月そのままになっていた。今年最後の研究者向けの記事として続編を書くことにする。
ミオサプレッシンのC末端はFLRF-アミドという構造であった。この構造は、二枚貝の神経節から心拍上昇活性をもつペプチドとして発見されたFMRF-アミドペプチドと似ている。C末端がFMRF-アミドと類似の構造をもつペプチドは様々な無脊椎動物からいくつも発見され、刺胞動物から昆虫まで多様な生理活性を有していることが知られている。これらペプチド類は、FMRF-アミド関連ペプチド(FaRP)と総称されている。
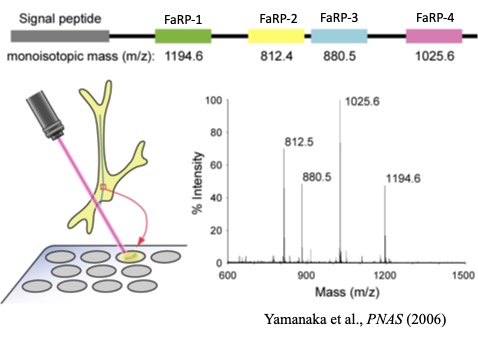
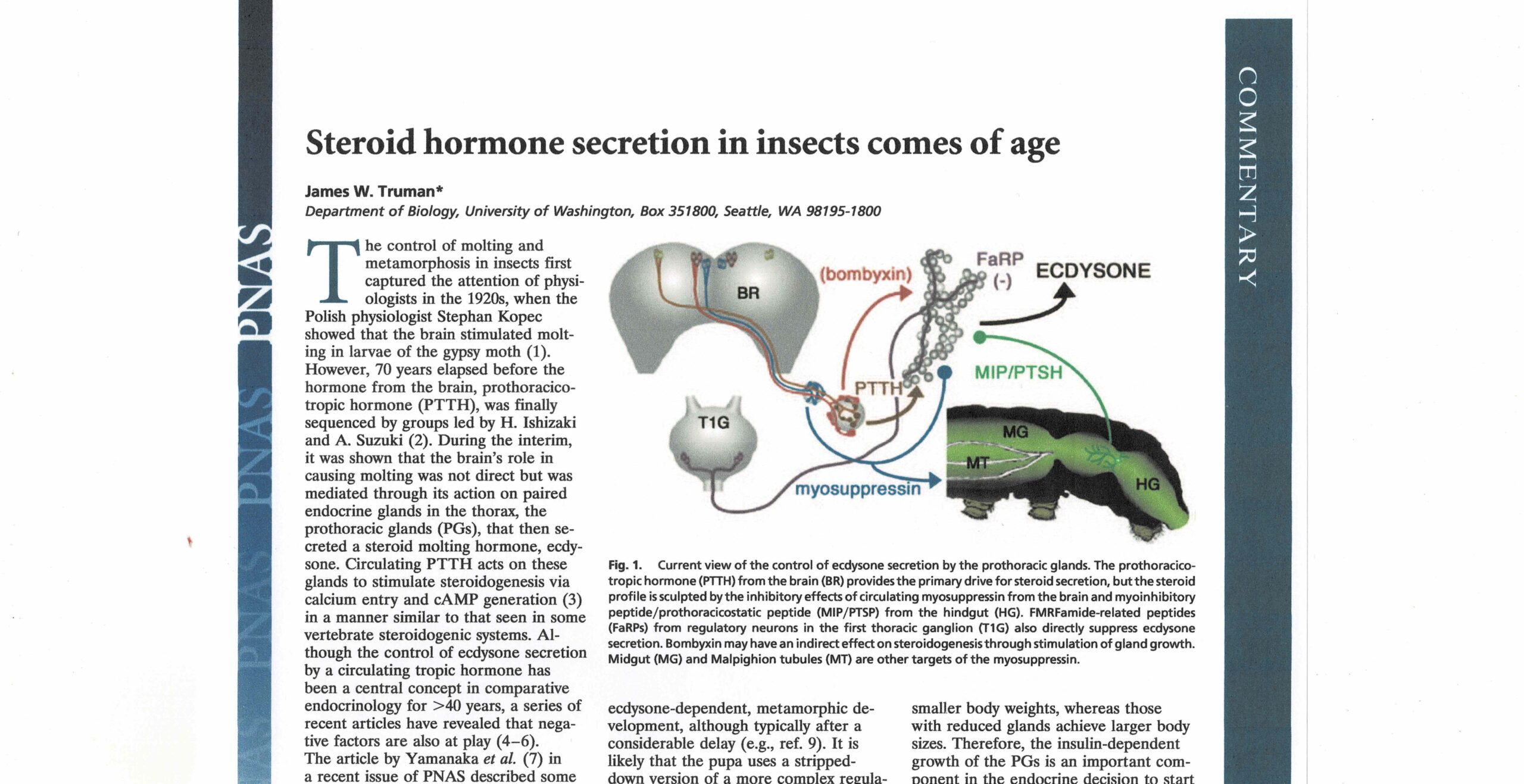
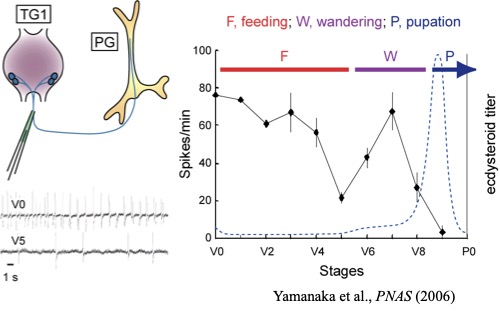
カイコミオサプレッシンの構造決定後、遺伝子のクローニングにも成功し、ミオサプレッシンの前駆体構造はショウジョウバエをはじめ昆虫間で保存性が高いことが分かった。いずれも、前駆体のC末端部分が成熟ペプチドであり、そのC末端に3残基の塩基性アミノ酸が続いていた。ところで、ショウジョウバエにはミオサプレッシン遺伝子以外のFaRP遺伝子が存在し、その遺伝子には13個のFaRPがコードされていることが知られていた。この類縁遺伝子と相同性がある遺伝子がカイコにも存在し、4種類のFaRPがコードされていることを明らかにできた。ミオサプレッシン遺伝子と比べると昆虫間での相同性はそれほど高くないが、複数のFaRPがコードされている点は同じである。また、カイコFaRPのうち3種類はRF-アミドを認識する抗体との反応を指標にペプチドとして単離することもできた。これら4種類のカイコFaRPを化学合成して、その前胸腺内cAMP抑制活性を調べたところ、いずれのペプチドもミオサプレッシンの1万倍以上の濃度でないと抑制活性を示さなかったことが分かった。そのような高濃度でしか活性を示さないことから血液を介したホルモンとして作用しているとは考えにくい。そんな時、共同研究者の田中良明さんから「RF-アミドを認識する抗体で前胸腺を染色すると、前胸腺の表面にある神経軸索が強く染まる」という情報をもらった。そこで、その神経軸先を取り出し、マトリックスを加えてMALDI TOF-MSを測定すると、4種類のFaRPの分子量に相当する分子イオンピークが全て観測された。つまり、この神経軸索を介して4種類のFaRPが前胸腺へ運ばれていると考えられる。また、免疫組織化学により、その神経軸先は胸部第1神経節にある2対の神経分泌細胞から伸びていることが分かった。さらに、電気生理学的手法で、この神経の発火頻度(神経分泌が盛んな時に発火頻度が高くなる)を終齢カイコの成長を追って調べると、脱皮直後の若い5齢幼虫では盛んに発火が見られるが、吐糸直前のワンダリング期と呼ばれる時期に発火頻度が下がり、その後少し上がり、蛹になる直前に急激に下がる(ほぼ発火しなくなる)ことが分かった。血液中のエクジソン濃度とミラー(逆相関)関係であった。つまり、この神経の分泌活動が前胸腺の分泌を抑えていて、エクジソン分泌が盛んになる時期には神経活動が抑えられていると考えられる。また、神経ペプチド(FaRP)が神経軸索を介して前胸腺に運ばれ、前胸腺細胞の表面で分泌されるために局所的には非常に高濃度になっていることが推測される。FaRPが高濃度でないと抑制活性を示さなかったことの説明になる。なお、私が大学院生の頃に、胸部神経節から前胸腺へ伸びる神経によって前胸腺が抑制されていることを、山口大学の岡島先生がヨトウガを用いた実験で示されていたと記憶している。ただし、その実体が神経ペプチドであるとは全く考えられていなかった。
このように、前胸腺のエクジソン合成・分泌は前胸腺刺激ホルモン(PTTH)が適切な時期に血液中に放出されることで促進され、脱皮や変態が誘導される。それ以外の時は血液中に分泌されているミオサプレッシンや、神経軸索を介して前胸腺近傍で分泌されるFaRPで抑制されていると考えられる。なお、促進性のPTTHと、抑制性のミオサプレッシンやFaRPを前胸腺に同時に作用させると、抑制されることが分かっている。つまり、昆虫にとって脱皮や変態は、そのタイミングがきちんとコントロールされていて、むやみやたらに脱皮・変態が起きないように二重三重の抑制機構によって制御されていることを示している。
以上の研究成果は、カリフォルニア大学リバーサイド校教授(現在)の山中直岐君が在学時に得たもので、国際的評価が高い雑誌PNASの表紙にもなり発表することができた、大学院5年間でミオサプッレッシン発見からここまでの成果をひとりで完成させた彼の能力の高さを感じた。また、PNASの次号で、昆虫生理学分野の権威であるワシントン大学のJ. W. Truman教授によりコメンタリー記事で研究内容が紹介された。当時、この分野で注目を受けた研究のひとつであった。
「ミオサプレッシン研究から派生した成果」のひとつ「蛹休眠」との関係についても紹介したいと思っていたが、少し長くなったので、年明けに「前胸腺抑制神経ペプチド(3)」を書くことにする。
遺伝子から予想される前駆体構造